the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 08 Jan 2019
| 08 Jan 2019
High-precision U–Pb ages in the early Tithonian to early Berriasian and implications for the numerical age of the Jurassic–Cretaceous boundary
Luis Lena
Rafael López-Martínez
Marina Lescano
Beatriz Aguire-Urreta
Andrea Concheyro
Verónica Vennari
Maximiliano Naipauer
Elias Samankassou
Márcio Pimentel
Victor A. Ramos
Urs Schaltegger
The numerical age of the Jurassic–Cretaceous boundary has been controversial and difficult to determine. In this study, we present high-precision U–Pb geochronological data around the Jurassic–Cretaceous boundary in two distinct sections from different sedimentary basins: the Las Loicas, Neuquén Basin, Argentina, and the Mazatepec, Sierra Madre Oriental, Mexico. These two sections contain primary and secondary fossiliferous markers for the boundary as well as interbedded volcanic ash horizons, allowing researchers to obtain new radioisotopic dates in the late Tithonian and early Berriasian. We also present the first age determinations in the early Tithonian and tentatively propose a minimum duration for the stage as a cross-check for our ages in the early Berriasian. Given our radioisotopic ages in the early Tithonian to early Berriasian, we discuss implications for the numerical age of the boundary.
- Article
(4756 KB) - Full-text XML
-
Supplement
(907 KB) - BibTeX
- EndNote





The age of the Jurassic–Cretaceous (J–K) boundary remains one of the last standing Phanerozoic system boundaries with a numerical age not tied by adequate radioisotopic data. The numerical division of the geological record is ultimately dependent on accurate and precise radioisotopic ages of well-defined fossiliferous datums. Over the years the numerical age of the J–K boundary has been difficult to measure due to the lack of datable horizons close to boundary markers, which made it difficult to ascribe a radioisotopic age directly to fossiliferous datums. Consequently, the ill-defined age of the boundary has led to widely variable timescales for the Late Jurassic to Early Cretaceous (Channell et al., 1995; Gradstein et al., 1995; Lowrie and Ogg, 1985; Malinverno et al., 2012; Ogg, 2012; Ogg et al., 1991; Ogg and Lowrie, 1986; Pálfy, 2008; Pálfy et al., 2000). These various approaches attempted to ascribe an age to the J–K boundary; nevertheless, the different estimates for the age of the boundary lacked reproducibility, varying from 135 to 145 Ma, with a high degree of uncertainty and very little overlap. The most recently used timescale of the Late Jurassic to Early Cretaceous is the M-sequence model of Ogg (2012). The model relies on the integration of data from a variety of fields such as M-sequence magnetic anomalies from the northwest Pacific Ocean, magnetostratigraphy, biostratigraphy, cyclostratigraphy, and scarce radioisotopic ages. The model is based on the marine magnetic anomalies timescale of the northwestern Pacific Ocean (Channell et al., 1995; Larson and Hilde, 1975; Tamaki and Larson, 1988). The interval encompasses ∼1000 km of oceanic crust over a period of ∼35 Myr in the northwestern Pacific. The age of the polarity changes in the northwestern Pacific was dated by key fossiliferous datums from Mediterranean Tethys sedimentary sequences via the correlation with magnetostratigraphy in these sequences (Grabowski, 2011; Kent and Gradstein, 1985; Ogg et al., 1991; Ogg and Lowrie, 1986). The duration of the magnetic reversal changes are provided by cyclostratigraphic studies (Huang et al., 2010a, b) for some of the magneto-zone intervals and thus used to calculate a decreasing spreading rate with the distance associated with the magneto-anomalies in the Hawaiian spreading center. The numerical age of stage boundaries from the Berriasian to Oxfordian were then back-calculated from the age of the M0r at the base of the Aptian. The age of the M0r used was 126.3±0.4 Ma, which is the combination of the cycle duration of the Albian stage (Huang et al., 2010a) tied to a U–Pb age from the Aptian–Albian boundary of 113.1±0.3 Ma (Selby et al., 2009). This linear fitting model is the basis for various Late Jurassic and Early Cretaceous stage boundary numerical ages.
In the specific case of the J–K boundary the projected age of the M-sequence age model was 145.0±0.8 Ma (Ogg, 2012), which is almost identical to the radioisotopic age reported in Mahoney et al. (2005) of 145.5±0.8 Ma (recalculated by Gradstein, 2012) for the sill intruded in Berriasian sediments in the Shatsky Rise with magnetization M21-M20. Furthermore, the magnetization of borehole 1213B is reasonably close to what has become a reliable secondary marker for the J–K boundary, the M19.2n (Wimbledon, 2017, and references therein). However, studies that obtained radioisotopic ages directly from sedimentary sequences that spanned the J–K boundary reveal much younger ages for the boundary (Bralower et al., 1990; López-Martínez et al., 2015; Vennari et al., 2014).
Recently, the base of the Calpionella Zone (the Alpina Subzone) has been selected as a principal biostratigraphic marker for the base of the Berriasian (Wimbledon, 2017). Nevertheless, its presence alone is not sufficient to locate the boundary, and secondary markers such as calcareous nannofossils and magnetostratigraphy are essential additional constraints to aid the definition of the boundary, with the latter allowing sections to be normalized against a global framework. The most complete studies of the J–K boundary from a biostratigraphical and magnetostratigraphic standpoint are located in Mediterranean Tethys. Nevertheless, the radiometric age of the boundary is poorly defined in the Mediterranean Tethys due to the absence of active volcanism close by during the time of deposition of these sedimentary sequences. In this way, the western Tethys (proto-Gulf of Mexico) and the Austral Basins (Neuquén Basin, Argentina) offer a good opportunity to advance the study of the radioisotopic age of the J–K boundary. Contrary to the Mediterranean Tethys, the sedimentary sequences in the proto-gulf and Austral realms were deposited close to active plate boundaries where significant volcanism took place, which enabled the deposition of datable horizons suitable for U–Pb geochronology. Recently, calpionellid biostratigraphy has been reported in both regions (López-Martínez et al., 2013b, 2017), opening possibilities for better correlations with the Mediterranean Tethys. It is worth noting that even if the calpionellid biostratigraphy of the Neuquén Basin is still not complete and global correlations are still tentative, for now they are the only known basins with occurrences of calpionellid as markers around the J–K boundary in the Austral realm with abundant datable horizons. A general definition of the J–K boundary would, however, need to be of global validity and allow for correlation with the Tethys realm.
In the present study, we date two independent sections, one in Mexico and one in Argentina, using precise radioisotopic geochronological methods. We present high-precision U–Pb age determinations using chemical abrasion–isotope dilution–thermal ionization mass spectrometry (CA-ID-TIMS) techniques to date zircon from interbedded volcanic ash layers in the Las Loicas section, Neuquén Basin, Argentina, and the Mazatepec section, Mexico. Such dates have proved to yield robust estimates for the timing of the stratigraphic record, especially in combination with Bayesian age–depth modeling (e.g., Ovtcharova et al., 2015; Baresel et al., 2017; Wotzlaw et al., 2017). The coupling of high-precision U–Pb geochronology and age–depth modeling allowed us to ascribe specific numerical ages to key taxa in the early Berriasian and late Tithonian in the studied sections. We also report new nannofossil data from the section in Mexico such as the first occurrence (FO) of Nannoconus steinmannii steinmannii and the FO of Nannoconus kamptneri minor (Fig. 2). Additionally, we also present the first radioisotopic age in the early Tithonian at the base of the Virgatosphinctes andesensis biozone in the La Yesera section, Neuquén Basin, close to the Kimmeridgian–Tithonian boundary (KmTB) (Riccardi, 2008, 2015; Vennari, 2016). Lastly, our geochronological data allow us to reevaluate the numerical age of the J–K boundary and discuss some complications with the currently accepted age of ∼145 Ma.
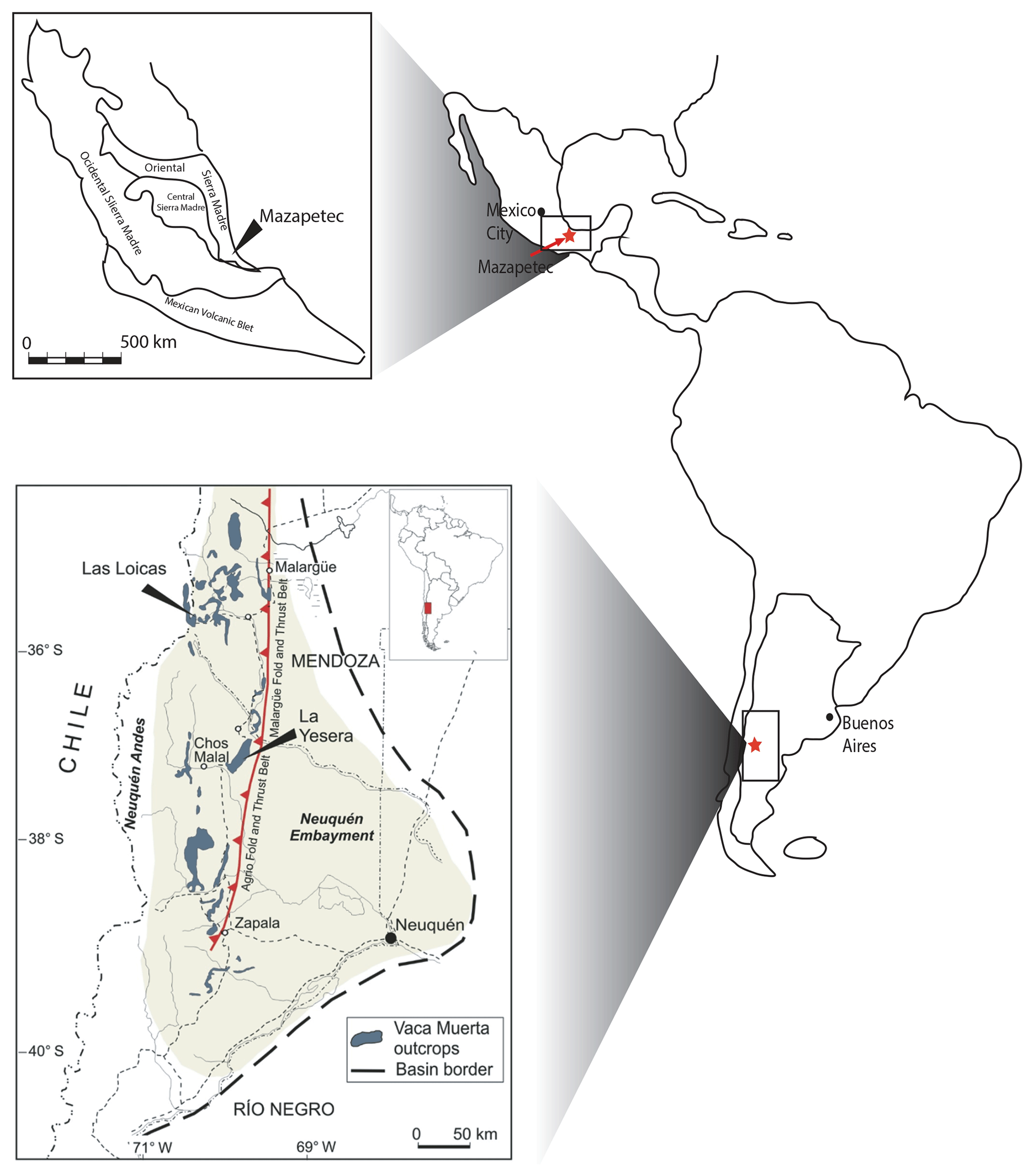
Figure 1Location of the studied sections and the general geological context of each section.
To investigate the numerical age of the J–K boundary, we have selected two sections where J–K boundary markers such as ammonites, calpionellids, and calcareous nannoplankton have been recognized. The first section is Las Loicas, exposed along the national road 145 (Argentina) from Bardas Blancas to the international border at the Pehuenche Pass. It is located near the Argentine–Chilean border, approximately 1 km to the southwest of the settlement Las Loicas (Fig. 1). Geologically, the Las Loicas section (Vennari et al., 2014) is located in the Vaca Muerta Formation, Neuquén Basin, Argentina (Fig. 1). The Neuquén Basin in western Argentina accumulated an almost continuous record of 7000 m of sediments from the Late Triassic to early Cenozoic. The basin is located on the eastern side of the Andes in Argentina between 32 and 40∘ S latitude (Fig. 1). The basin has a triangular shape, covers an area of over 12002 km, and is bounded to the west by the Andean magmatic arc on the active margin of the South American Plate, to the northeast by the San Rafael Block, and to the southeast by the North Patagonia Massif (Fig. 1). Two main regions are commonly recognized in the basin: the Neuquén Andes to the west and the Neuquén Embayment to the east (Fig. 1). The Neuquén Embayment is relatively undeformed, in contrast to the Neuquén Andes where the late Cretaceous–Cenozoic deformation has resulted in the development of a series of N–S-oriented fold and thrust belts: Aconcagua, Malargüe, and Agrio, where a substantial part of the Mesozoic sequence outcrops (Legarreta and Uliana, 1991, 1996).
The Vaca Muerta Fm. is a 217 m thick sedimentary sequence of marine shales and limestones, which spans an interval from the lower Tithonian (Virgatosphinctes andesensis biozone) to the upper Berriasian (Spiticeras damesi biozone) (Aguirre-Urreta et al., 2005; Kietzmann et al., 2016; Riccardi, 2008, 2015). In Las Loicas, the Substeueroceras koeneni and Argentiniceras noduliferum ammonite biozone and calcareous nannofossils have been described by Vennari et al. (2014). Recently, López-Martínez et al. (2017) reported the occurrence of upper Tithonian to lower Berriasian calpionellids, which is the only known section where the primary markers for the J–K boundary occur together in the Argentinian Andes. The section contains several ash beds, which allowed for precise age bracketing of the boundary using high-precision U–Pb geochronology.
The La Yesera Section is located 50 km north of the town of Chos Malal in the northern sector the Neuquén Basin (Fig. 1) and is exposed along the national road 40. Geologically, the La Yesera section (Fig. 2c) represents a distal portion of the basin farther from the magmatic arc than the Las Loicas section. Tuff beds are less frequent than in the Las Loicas section and generally thinner. The section has a total thickness over 400 m and is one of the best continuous exposures of Tithonian ammonite zones, from the early Tithonian Virgatosphinctes mendozanus to the Neocomites wichmanni (early Valanginian; Aguirre-Urreta et al., 2014). The section also has one of the best-exposed contacts between the Vaca Muerta Fm. and the Tordillo Fm.
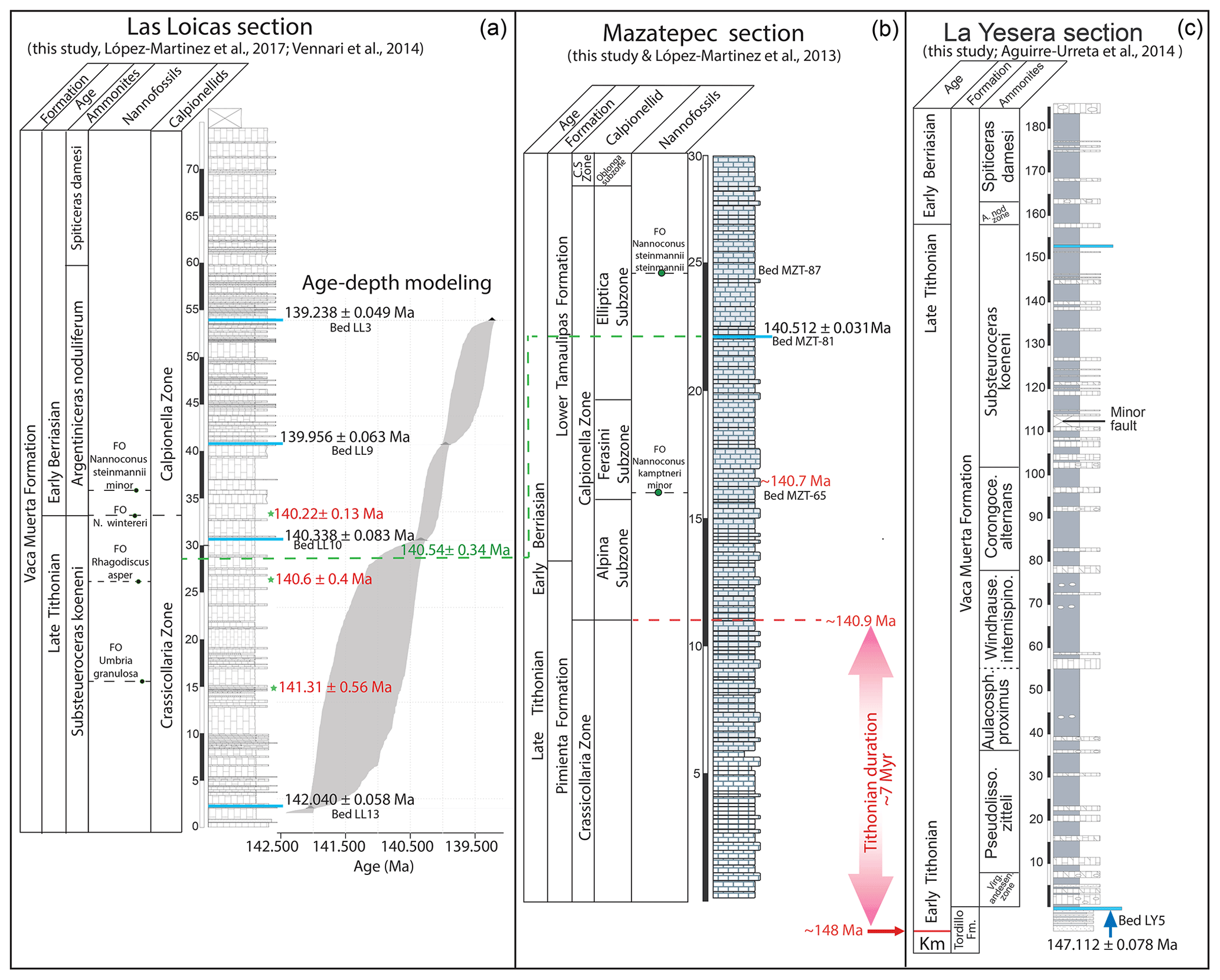
Figure 2Age correlation between the Las Loicas, Mazatepec, and La Yesera sections. (a) Las Loicas section: ash beds in light blue with respective name and U–Pb dates in black font; age–depth modeling ages are in red font next to green stars (this study); ammonite and nannofossil zonation after Vennari et al. (2014); calpionellid zonation after Lopez-Martinez et al. (2017). (b) Mazatepec section: ash bed in light blue with respective name and U–Pb age in black font, age calculated from sedimentation rate in red font (this study); calcareous nannofossils (this study); calpionellid zonation after Lopez-Martinez et al. (2013b). (c) La Yesera section: ash bed in light blue with U–Pb age (Aguirre-Urreta et al., 2014).
The Mazatepec section is located in the Puebla State, Mexico, southeast of Mexico City. Geologically, the Mazatepec section exposes the Pimienta and the lower Tamaulipas formations of the Sierra Madre Oriental geological province, Mexico (Fig. 1). The Sierra Madre Oriental is one of the many tectonic terranes composed of Mesozoic volcano-sedimentary sequences deformed during the Late Cretaceous and early Cenozoic during the Laramide Orogeny in Mexico (Campa and Coney, 1983; Suter, 1980). A rift sequence characterizes the tectonic evolution of the proto-gulf in the Late Triassic–Oxfordian due to the rifting of Pangea characterized by continental sedimentation controlled by narrow grabens with no marine sedimentation taking place (Salvador, 1987). The post-rift phase is characterized by ample marine carbonate platforms of shallow waters. During the Tithonian to Early Cretaceous, stable tectonic and climatic conditions prevailed with the sedimentation being significantly slower with the development of shallow marine water sedimentation, namely the deposition of the Pimienta Fm. (carbonates) and Tamaulipas Fm. (argillaceous limestones, shales) (Padilla and Sánchez, 2007). The Pimienta Fm. is composed of darkish clayey limestones and the Tamaulipas Fm. is a gray limestone (López-Martínez et al., 2013b; Suter, 1980). The section has a dense occurrence of late Tithonian Crassicollaria Zone (Colomi Subzone) and early Berriasian calpionellids from the Calpionella Zone (Alpina, Ferasini, and Elliptica subzones) to the Calpionellopsis Zone (Oblonga Subzone). In the upper part of the section, ash beds are scarce and occur at distinct levels. Ash bed MZT-81 is situated within the Elliptica Subzone in the lower Tamaulipas Formation (Fig. 2b).

Figure 3(a–h) Representative calcareous nannofossils from the Mazatepec section, Mexico. (a) Conusphaera mexicana Trejo (BAFC-NP 4190) (2 m), (b) Conusphaera mexicana Trejo (BAFC-NP 4196) (11 m), (c) Hexalithus noeliae Loeblich and Tappan (BAFC-NP 4195) (7.5 m), (d) Hexalithus geometricus Casellato (BAFC-NP 4205) (25 m), (e) Nannoconus kamptneri minor Bralower (BAFC-NP 4201) (16 m), (f) Nannoconus globulus Brönnimann (BAFC-NP 4205) (25 m), (g–h) Nannoconus steinmannii subsp. steinmannii Kamptner (BAFC-NP 4205) (25 m). Our suggestion is to eliminate calcareous nannofossil images published previously from the Las Loicas section in order to avoid more confusion with taxonomy.
The nannofossil biostratigraphy for the Mazatepec section was based on 17 samples from the Pimienta and Tamaulipas formations. For detailed calcareous nannofossil examination, simple smear slides were prepared using standard procedures (Edwards, 1963). Observations were made and photographs were taken using a polarizing microscope Leica DMLP increased 1000X and accessories such as λ plaster and a blue filter. The slides are deposited in the Repository of Paleontology, Department of Geological Sciences, University of Buenos Aires, under catalog labels BAFC-NP: nos. 4190–4206. Photomicrographs of selected species are shown in Fig. 3; the distribution chart for the calcareous nannofossil species is presented in Supplement Fig. S1.
We have used U–Pb zircon CA-ID-TIMS dating techniques for single zircon grains, which yields 206Pb∕238U dates at 0.1 %–0.05 % precision. The depositional age of ash beds has been calculated from the weighted means of the four youngest overlapping 206Pb∕238U dates (Fig. 4), assuming that older grains record prolonged residence of zircon in the magmatic systems as well as intramagmatic recycling. In the text, all quoted ages of ash beds are weighted mean 206Pb∕238U ages corrected for initial 230Th disequilibrium.
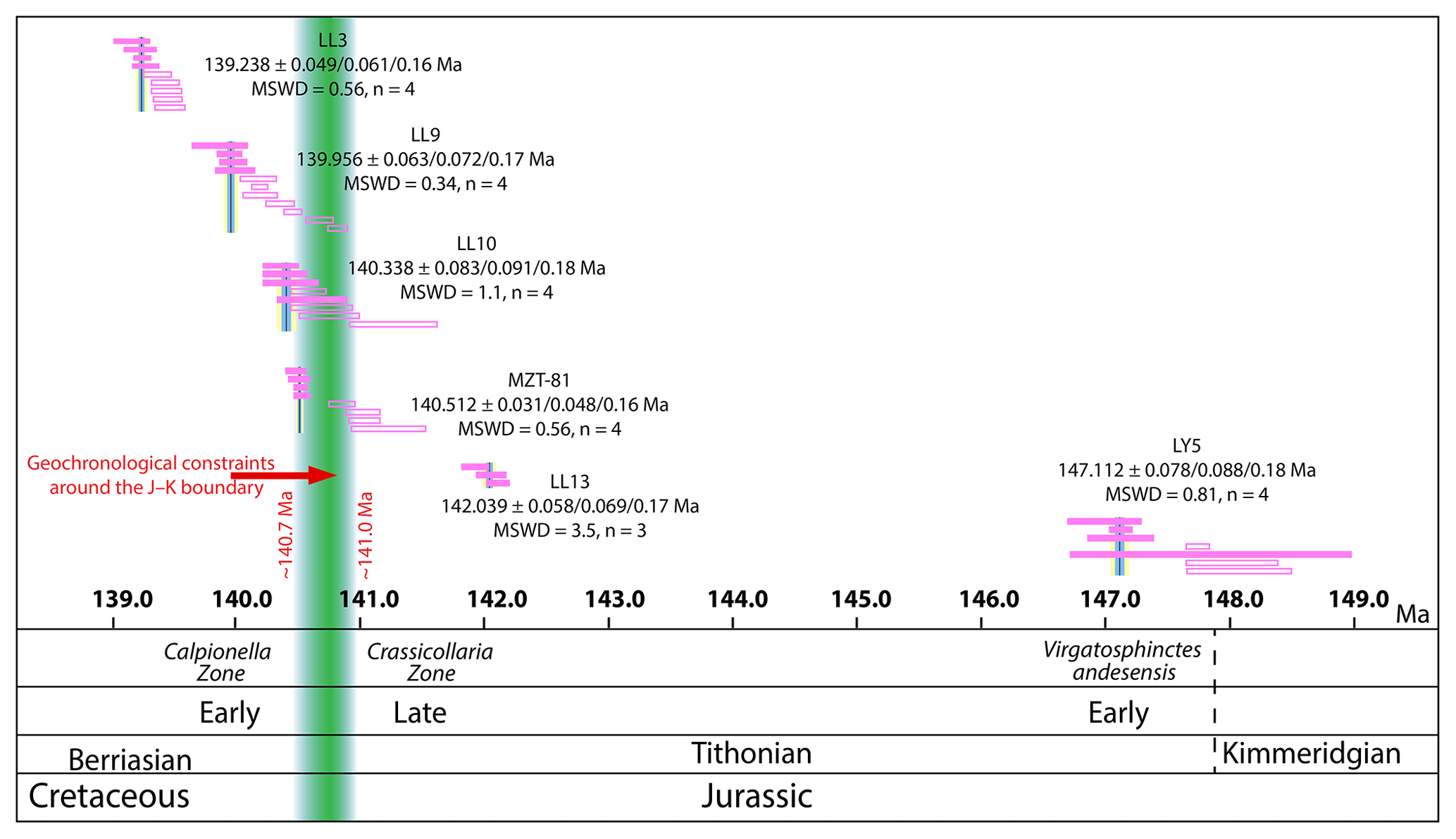
Figure 4U–Pb-weighted mean ages of the dated ash beds and the ages and projected ages of the J–K boundary interval, the base of the Calpionella alpina zone, the top of the Crassicollaria Zone, the Virgatosphinctes andesesis zone, and the KmTB at ∼148 Ma. Color bars represent grains considered in the weighted mean age.
The ages of the various paleontological markers in Las Loicas have been calculated using the Bayesian age–depth model Bchron of Haslett and Parnell (2008) and Parnell et al. (2008). The model outputs an uncertainty envelope, which is presented in Fig. 2b. The age–depth results are reported in TS.2, with age assigned to every meter of stratigraphic height. The Bchron code used in the R statistical package environment (R Core Team, 2013) is included in Supplement Sect. S6.
4.1 Calcareous nannofossil biostratigraphy in Mazatepec
Eighteen nannofossil species have been recognized in Mazatepec (Fig. S1). The heterococcoliths are mostly represented by Watznaueriaceae including Watznaueria barnesae, W. britannica, W. manivitae, Cyclagelosphaera margerelii, and C. deflandrei; Zeugrhabdotus embergeri is another frequent constituent. The nannoliths are represented by Conusphaera mexicana, Polycostella senaria, Hexalithus noeliae, Nannoconus globulus, and N. kamptneri minor. These nannofossils indicate a late Tithonian to early Berriasian age for the Pimienta Formation and the lower part of the Tamaulipas Formation. The assemblage composed of Conusphaera mexicana, Polycostella scenario, and Hexalithus noeliae indicates a late Tithonian age. The only useful biological event recognized is the FO of N. kamptneri minor. An increase in the diversity of nannofossils is identified with 11 species, among which the presence of N. steinmannii steinmannii stands out (Fig. 2b).
4.2 U–Pb geochronology, age interpretations, age–depth modeling
A total of six ash beds were dated: four in the Las Loicas section, one in the Mazatepec section, and one in the La Yesera section. In the Las Loicas section, LL3 yielded an age of , LL9 , LL10 , and LL13 an age of Ma. In La Yesera, ash bed LY5 yielded an age of Ma, and in Mazatepec MZT-81 yielded an age of Ma (Fig. 4). All zircons considered in the age distribution of the ash are interpreted from ashfall deposits from nearby volcanic eruptions. The final weighted mean ages are interpreted as a depositional age for each ash bed. Uncertainties are reported as where X includes analytical uncertainty, Y includes additional tracer (ET2535) calibration uncertainty, and Z includes additional 238U decay constant uncertainty. A full and detailed description of the techniques, sample preparation, laboratory procedures, data acquisition, and data treatment is provided in the Supplement. The full U–Pb data set is reported in Table S1 in the Supplement. Age–depth statistical modeling was performed by outputting a numerical age for every meter of the Las Loicas sections, with a 95 % confidence precision interval. The results with a meter-by-meter resolution are reported in Table TS.2.
4.3 Numerical age of faunal assemblages in studied sections
In Fig. 2a, the various markers and assemblages are indicated, as are the ages of the ash beds. In Las Loicas, López-Martínez et al. (2017) reported on the late Tithonian Crassicollaria Zone and the Colomi Subzone (upper Tithonian) based on the occurrence of Calpionella alpina Lorenz, Crassicollaria colomi Doben, Crassicollaria parvula Remane, Crassicollaria massutiniana (Colom), Crassicollaria brevis Remane, Tintinnopsella remanei (Borza) and Tintinnopsella carpathica (Murgeanu and Filipescu), and the FO of the Umbria granulosa granulosa and Substeueroceras koeneni ammonite zone (Vennari et al., 2014). Our Bchron model age predicts an age of 141.31±0.56 Ma for the faunal assemblage of Crassicollaria parvula and Crassicollaria colomi and the FO of Umbria granulosa granulosa Fig. 2b). Another late Tithonian marker in Las Loicas is the FO of Rhagodiscus asper, also within the Crassicollaria Zone, with a Bchron age of 140.60± Ma (Fig. 2a).
In Las Loicas some early Berriasian markers are present. For instance, the FOs of Nannoconus kamptneri minor (Figs. 2a, S1) and Nannoconus steinmannii minor are considered indicators of the early Berriasian (Vennari et al., 2014). Here they overlap with the base of the Argentiniceras noduliferum ammonite zone (López-Martínez et al., 2017; Vennari et al., 2014). The occurrence of the acme of Calpionella alpina (small and spherical) and scarce specimens of Crassicollaria massutiniana, Tintinnopsella remanei, and T. carpathica suggest an early Berriasian age (López-Martínez et al., 2017) (Fig. 2a). These assemblages are bracketed by ash beds LL9 (139.956±0.063 Ma) and LL10 (140.338±0.083 Ma) (Fig. 2a) and overlap with the FO of Nannoconus kampteri minor and Nannoconus steinmannii minor, the base of the Argentiniceras noduliferum zone, and the base of the Alpina Subzone (ca. 34 m of stratigraphic height) (Fig. 2a). The Bchron model age for this assemblage is 140.22±0.13 Ma (Fig. 2a).
In Mazatepec, ash bed MZT-81 is located within the Elliptica Subzone and has an age of 140.512±0.031 Ma. (Fig. 4). Due to the lack of datable horizons close to the Alpina Subzone in Mazatepec we have resorted to assumed sedimentation rates to back-calculate the age of the base of the Alpina Subzone. Here we assume the sedimentation rate to be 2.5 cm ka−1. Although there are no data on actual sedimentation rates in the Pimienta Fm., this rate is realistic for similar coeval deposits (e.g., Grabowski et al., 2011) as well as with the tectonic and environmental stability of the Sierra Madre Oriental in the Tithonian–Berriasian stages (Padilla and Sánchez, 2007). It is worth noting that our new data allow only a confident numerical age for the Elliptica Subzone (Fig. 5).
Ash bed LY-5 was located below the contact, and it yielded an age of 147.112±0.078 Ma (Fig. 2C). The ash bed is located in the Tordillo Fm., 1.5 m below the contact with the Vaca Muerta Formation and thus very close to the base of the Virgatosphinctes andesensis zone.
5.1 The chronostratigraphic and biostratigraphic framework of the studied sections
In the past decade significant strides have been made in fixing the J–K boundary by coupling calpionellids, calcareous nannofossils, ammonites, and magnetostratigraphy (Wimbledon, 2017; Wimbledon et al., 2011). Correlations between sections within the Mediterranean Tethys have become consistent to the point of a trustworthy correlation framework being developed for the various markers (calpionellids, nannofossils, ammonites, and magnetostratigraphy) of the J–K boundary (Wimbledon, 2017, references therein). Even though important biostratigraphic studies have been carried out in other regions outside of the Mediterranean Tethys, such as in the proto-gulf and the Argentinian Andes, the correlation between these regions remains uncertain. Notably, the lack of magnetostratigraphic data in studies from the proto-gulf (López-Martínez et al., 2013a, b) and the Argentinian Andes (López-Martínez et al., 2017; Vennari et al., 2014) is a challenge and leaves room for ambiguity in biochronostratigraphical correlations. Here we attempt to describe the limitations of the biostratigraphical markers in the studied sections.
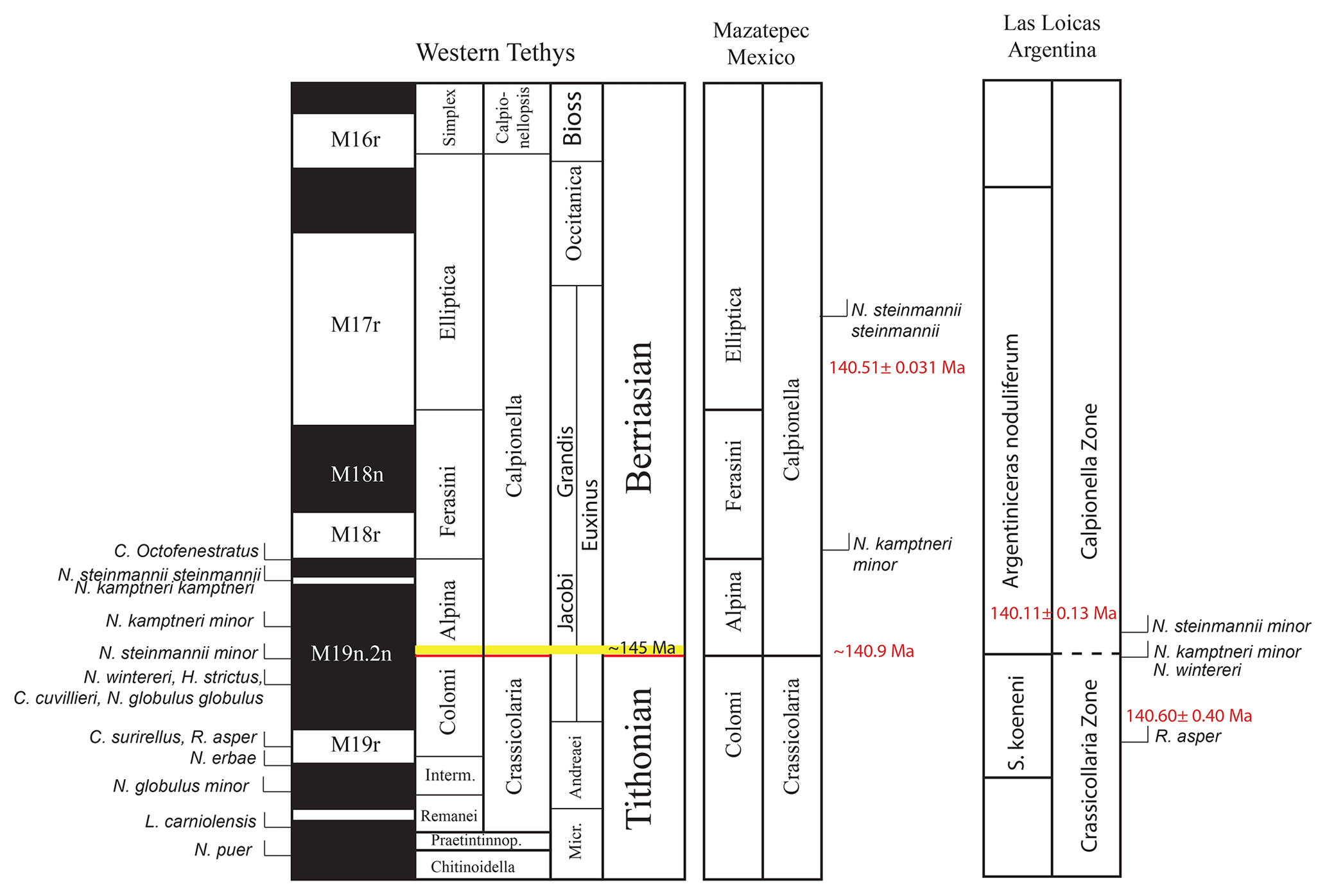
Figure 5Tentative correlation of the studied sections with the western Tethys correlation scheme of Wimbledon et al. (2017).
In Mazatepec, only two important calcareous nannofossil bioevents are recognized, i.e., the FO of N. kamptneri minor and N. steinmannii steinmannii. In the Tethys realm, the former bioevent occurs within the M19.2n, slightly above the base of the Alpina Subzone (Bakhmutov et al., 2018), and it is used as an upper limit to the base of the Alpina Subzone (Wimbledon et al., 2013). In Mazatepec, the FO of N. kamptneri minor occurs 5 m above the base of the Alpina Subzone; however, it is within the lower Ferasini Subzone and thus slightly younger than in the Mediterranean Tethys. Another bioevent in Mazatepec is the FO of N. steinmannii steinmannii, which occurs within the Elliptica Subzone. This marker has been shown in the past to occur within the Elliptica Subzone and coincident within the M17r (Casellato, 2010), but has been found as low as the Alpina Subzone, the base of M18r (Bakhmutov et al., 2018; Hoedemaeker et al., 2016; Lukeneder et al., 2010), or even lower (Svobodová and Košťák, 2016). Even though our new calcareous nannofossils from Mazatepec are an addition to the biostratigraphic framework of the sections, this is very preliminary and does not provide any definite constraints for the J–K boundary or the base to the Alpina Subzone in the section. Valuable markers such as N. steinmannii minor, N. wintereri, and H. strictus have not yet been reported. Furthermore, no calcareous nannofossils have been reported below the base of the Alpina Subzone in Mazatepec. Nevertheless, we feel that the FO of N. kamptneri minor so close to the base of the Alpina Subzone in Mazatepec provides confidence for futures studies in the section.
In the Mediterranean Tethys, important markers for the J–K boundary are the first appearance datum (FAD) of N. kamptneri minor and N. wintereri. In the Tethys, these two markers usually occur in the middle of the M19.2n, but in distinct stratigraphic horizons and commonly bracketing the base of the Alpina Subzone (Wimbledon, 2017; Wimbledon et al., 2013). N. wintereri, for instance, occurs below the base of the Alpina Subzone (Elbra et al., 2018; Svobodová and Košťák, 2016; Wimbledon et al., 2013) and in one occurrence as low as the M19r (Lukeneder et al., 2010). In Las Loicas, on the other hand, both occur virtually within the same stratigraphic range (Vennari et al., 2014). The close FO of N. kamptneri minor, N. wintereri, C. deflandrei, and M. pemmatoide in Las Loicas (Vennari et al., 2014) is also troublesome.
The most important secondary marker for the J–K boundary is the FAD of N. steinmannii minor, which usually occurs in the vicinity of the Alpina Subzone (Wimbledon, 2017), below it (Bakhmutov et al., 2018), and slightly above it (Hoedemaeker et al., 2016; Svobodová and Košťák, 2016). In Las Loicas, the FO of N. steinmannii minor is present and occurs in the vicinity of the Alpina Subzone; however, it is limited to a single sample (Vennari et al., 2014) and not continuous. Furthermore, in Las Loicas the FOs of N. kamptneri minor and N. wintereri are recorded below the FO of N. steinmannii minor. This order of occurrence in Las Loicas is contradictory because the FO of N. steinmannii minor is considered older than the FO of N. kamptneri minor and younger than the FO of N. wintereri. These circumstances suggest that condensation and/or preservation issues might be affecting the completeness and continuity of the calcareous nannofossil biostratigraphy in Las Loicas, thus impeding a reliable correlation between the Argentinian Andes and the Tethys.
Another possible issue with the biostratigraphy in Las Loicas pertains to a couple of calpionellid assemblages that might seem unusual when compared to the Mediterranean Tethys. The first is the presence of Tintinnopsella remanei in the upper part of the Crassicollaria Zone. This is a nontypical appearance in the Mediterranean Tethys, but it is usual in the western Tethys as discussed in López-Martínez et al. (2017). The second is the record of Crassicollaria massutiniana in the lowermost part of the Alpina Subzone. Even when it can be unusual, the presence of this species in the lowermost Berriasian does not affect the biozonation scheme as the Alpina Subzone is defined by the acme of Calpionella alpina in small and globular form and not the last occurrence (LO) of any species. Therefore, the Alpina Subzone is defined in Las Loicas in the same way as in the Mediterranean Tethys and can be used as a reasonable marker for the base of the Berriasian in Las Loicas.
In conclusion, there is still ambiguity in the biostratigraphic framework of the studied sections with regards to the J–K boundary markers. The incompleteness and frequency of key taxa call for further investigation and improvements to the biostratigraphy; important elements are still lacking for a definite and precise definition of the J–K boundary in both sections and correlations are still troublesome.
5.2 Constraining the numerical age of the J–K boundary between the studied sections
In Mazatepec, the middle of the Elliptica Subzone has an age of 140.512±0.031 Ma and consequently a numerical age in the lower Berriasian (Figs. 2 and 4). Conversely, in Las Loicas, the Bchron age model predicts that approximately the same age, i.e., 140.54±0.37 Ma (ca. 28.5 m; see TS.2), is found in the Crassicollaria Zone, 1 m above the FO of R. asper and thus late Tithonian (Fig. 2a). In other words, the age of ∼140.5 Ma in one section is coincident with late Tithonian fauna, and in the other it yields an age coincident with early Berriasian fauna. We see no reason to question the accuracy of the radioisotopic dates. It thus becomes apparent that both sections are offset in age, and Mazatepec is older than Las Loicas. Therefore, our geochronology points to limitations in the biochronostratigraphical correlation of these two sections.
Given the limitations of the biostratigraphy around the J–K boundary in both sections, our ability to quote a single numerical age for the J–K boundary is strongly hindered. Nevertheless, we feel that constraining, bracketing, and/or creating an age confidence interval for the J–K boundary using the biostratigraphical and geochronological constraints from both sections is a reasonable alternative to circumventing these limitations. To constrain the interval, we have tentatively chosen upper and lower limits to the interval based on the available biostratigraphic markers and their estimated ages that best bracket the J–K boundary. In Mazatepec, we suggest the FO of N. kamptneri as the upper biostratigraphical marker for the J–K age interval. In this section, the FO of N. kamptneri is close to the base of the Ferasini Subzone, and thus a subzone normally associated with the upper Alpina Subzone (Wimbledon, 2017, and references therein), the base of the 18r (Casellato, 2010), and M19n (Wimbledon et al., 2013), although it has recently been shown to be found at the base of the M19.2n (Bakhmutov et al., 2018). We feel that this could be used as a very conservative upper limit for the age of the J–K boundary. Using the sedimentation rate of 2.5 cm ka−1 in Mazatepec, we estimate the age of the FO of N. kamptneri and conceivably the base of the Ferasini Subzone to be ∼140.7 Ma (Fig. 2b). This is a conservative estimate for the upper age of the J–K boundary in Mazatepec and could very likely be older since the FO of N. kamptneri is commonly older than the base of the Ferasini Subzone (Wimbledon, 2017, and references therein). The base of the Alpina Subzone in Mazatepec is estimated to be ∼140.9 Ma, although a bracketing of the Alpina Subzone was not possible due to the absence of calcareous nannofossils commonly occurring at the base of the Alpina Subzone, such as N. steinmannii minor, or older diagnostic markers such as R. asper, N. erbae, and N. globulus. Therefore, a lower limit to the boundary in Mazatepec cannot be delineated.
Conversely, in Las Loicas, a few late Tithonian calcareous nannofossils occur in an assemblage with late Tithonian calpionellids such as the FO of R. asper, which is within the upper Crassicollaria Zone, and close to the FO of U. granulosa (Bralower et al., 1989; Casellato, 2010). These markers in Las Loicas allow for a lower age limit for the J–K boundary. Given these circumstances we suggest 1 m above the FO of R. asper as the lower limit of the J–K interval in Las Loicas. The Bchron model provides an age for the FO of R. asper at 140.60±0.4 Ma (ca. ∼27 m; see TS.2), which allows for a small overlap between the estimated age of the base of the Alpina Subzone in Mazatepec and the late Tithonian and early Berriasian assemblages in Las Loicas.
In summary, we have attempted to constrain the age of the J–K boundary using the biostratigraphical markers and our geochronology from Las Loicas and Mazatepec. Ash bed MZT-81 (middle of the Eliptica Subzone) suggests a minimum age. As a result, the age of the J–K boundary has to be older than 140.512±0.031 Ma, most likely older than ∼140.7 Ma (FO of N. kamptneri and/or the base of the Ferasini Subzone in Fig. 5; base of the M18r, possibly within M19.2n), but the latter age estimate derives from an approximate sedimentation rate (2.5 cm ka−1) that carries some uncertainty. In Las Loicas, the Bchron model age of the FO of R. asper (possibly the middle of the M19r) suggests a maximum age of the J–K boundary at 140.60±0.4 Ma. Given that the age of the Alpina Subzone in Mazatepec is estimated at ∼140.9 Ma, we suggest that the age of the J–K boundary be bracketed between 140.7 and 141.0 Ma. This interval accounts for the age of the boundary being slightly older than the base of the Alpina Subzone in Mazatepec due to the lack of secondary markers below the subzone. Our attempt to constrain the age of the J–K boundary is based only on the diagnostic markers for the boundary reported in the studied sections and the additional fact that we can calculate and estimate their ages, even if the chosen upper and lower limit of the interval has been proven to lie distant to the J–K boundary. Given the inherited uncertainties of the biostratigraphy and geochronology, we consider this age bracket as our best estimate for the J–K boundary interval.
5.3 The early Tithonian and the base of the Vaca Muerta Formation
The base of the Vaca Muerta Formation contains an early Tithonian ammonite assemblage of the Virgatosphinctes andesensis zone (Riccardi, 2008, 2015; Vennari, 2016). The gradational contact between the Vaca Muerta and the Tordillo formations is very well exposed in the La Yesera section and contains ash beds very close to the contact (Fig. S2b). We dated ash bed LY-5, and it yielded an age of 147.112±0.078 Ma (Fig. 2c). The ash bed is located in the Tordillo Fm., 1.5 m below the contact with the Vaca Muerta Formation and thus very close to the base of the Virgatosphinctes andesensis zone. This biozone is mostly equivalent to the Darwini Zone of the Tethys Ocean, which is broadly regarded as early Tithonian and widely distributed in various other regions including Mexico and Tibet (Riccardi, 2008, 2015; see Vennari, 2016, for a thorough review of the subject). Consequently, we suggest that the age of ash bed LY-5 (147.112±0.078 Ma) can be regarded as an age in the early Tithonian. This result is in good agreement with other studies that have dated the early Tithonian. For instance, Malinverno et al. (2012) quote an age of 147.95±1.95 Ma for the M22An magneto-zone, and Muttoni et al. (2018) suggest that the base of the Tethyan Tithonian (top Kimmeridgian) falls in the lower part of M22n with an age of ∼146.5 Ma.
Assuming the age of ash bed LY-5 (147.112±0.078 Ma) in La Yesera to be early Tithonian and coupling it with the estimated bracketed age of the J–K boundary (140.7–141. Ma), we can calculate a minimum duration for the Tithonian of ∼6–7 Myr (Fig. 2c). This is in good agreement with the current full duration of the Tithonian estimated at ∼7 Myr (145.5 to 152.1 Ma; see Ogg et al., 2016b). Furthermore, the M-sequence geomagnetic polarity timescale (MHTC12) of Malinverno et al. (2012) suggests a duration for the Tithonian of 5.75±2.47 Myr (i.e., between magneto-zones M22An and M19n.2n). Therefore, our new ages around the base of the Berriasian and close to the earliest Tithonian are in good agreement with other independent timescale estimates for the duration of the Tithonian. Incidentally, this result also has direct implications for the age of the KmTB. Currently, the age of the KmTB is 152.1±0.9 Ma according to the International Commission on Stratigraphy (ICS) (see also Ogg et al., 2016b). Admittedly, ash bed LY-5 is not at the KmTB, although it is close; therefore, we acknowledge that the age of the KmTB would have to be older than bed LY-5. Nevertheless, if the age of the KmTB is 152.1 Ma, it would imply that the Virgatosphinctes ammonite zone itself lasts more than ∼5 Myr, resulting in a total duration of ∼12 Myr for the Tithonian. It appears reasonable that our results for the early Tithonian are in agreement with other studies that dated the KmTB, and this also suggests that the current ICS KmTB age may need revision.
5.4 Implications for the numerical age of the J–K boundary
As of now, the age of the J–K according to the ICS is ∼145 Ma, which is ∼4 Myr older than our ages around the J–K boundary (Figs. 4 and 5). As we have explored in previous sections, the level of detail of the biostratigraphy in the studied sections needs improvement and fails to provide a precise constraint for the J–K boundary. A significant offset of potentially ∼600 kyr outlines the limitations of correlating biostratigraphy and geochronology between the two sections. Nevertheless, the disparity between our ages presented here and the current age of the J–K boundary is such that even with the biostratigraphical limitations and the absence of magnetostratigraphy, it calls for further attention to the numerical age of the J–K boundary. For instance, in Las Loicas the assemblage of Crassicollaria parvula and Crassicollaria colomi and the FO of Umbria granulosa granulosa have an age of 141.31±0.56 Ma (Fig. 2a), with the FO of R. asper at 140.60±0.4 Ma, which can be considered to lie within the late Tithonian and thus constrain the approximate age of the boundary. Furthermore, our age in the Elliptica Subzone in Mazatepec is at 140.512±0.031 Ma (early Berriasian). Worthy of attention is the age of ash bed LY5 in the Virgatosphinctes andesensis biozone (early Tithonian) at 147.112±0.078 Ma. These geochronological constraints make it fairly difficult to reconcile the base of the Berriasian to be ∼145 Ma and also has important implications for the duration of the Tithonian (see discussion above on the early Tithonian). From our new geochronological data, ∼145 Ma would most likely be an age in the middle of the Tithonian rather than the base of the Berriasian (Fig. 4). Other recent geochronological studies on the age of the J–K boundary using different geochronological approaches (e.g., Re–Os isochron ages from shales or LA-ICP-MS U–Pb ages on zircons) and in the Early Cretaceous are also at odds with the current age of the boundary. López-Martínez et al. (2015, 2017), Pálfy et al. (2000), and Tripathy et al. (2018) have published geochronological results that overlap within uncertainty with our age estimate of the J–K boundary (around 140–141 Ma). In summary, there is growing evidence that the age of the J–K boundary is most likely younger, although unequivocal evidence is still lacking.
The endurance of the numerical age of the boundary is mainly due to the perfect overlap between the M-sequence age model of Ogg (2012) and Mahoney et al. (2005). The latter authors dated a basaltic intrusion in lower Cretaceous (NK1) sedimentary rocks and argued that the age of the basalt would be close to the age of the J–K. Their age for the intruded basalt is 144.2±2.6 Ma (40Ar∕39Ar). This age was later corrected by Gradstein (2012) and Ogg et al. (2012) to 145.5±0.8 Ma with the recalibrated 40K decay constant of Renne et al. (2010). The magnetization of drill core 1213B proved to be between anomalies M19 and M20 (Sager, 2005), which was consistent at that time with the working model for the base of the Berriasian placed between M19 and M18 (now more precisely calibrated in the middle of the M19.2n; Wimbledon, 2017). This overlap was also in agreement with the numerical timescale of Gradstein et al. (1995). These facts have mainly been the anchors for the numerical age of the J–K boundary in the past years. However, analytical and biostratigraphical issues potentially reveal some inconsistencies in the numerical age for the boundary in Mahoney et al. (2005). For instance, the biostratigraphy of drill core 1213B poses problems. Bown (2005) pointed out that the sediments of this core are devoid of age-diagnostic NK1 nannofossils such as Conusphaera and Nannoconus. Important markers such as the family Cretarhabdaceae are present but in rare occurrences. Drill core 1213B is limited to the occurrence of nannofossils considered secondary markers and lacked any primary markers for the boundary. Even with the existing problems in the biostratigraphy of drill core 1213B, the magnetization of the dated basalt is in reasonable agreement with the magnetic timescale for the base of the Berriasian (Wimbledon, 2017). More importantly, it is worth pointing out that Mahoney et al. (2005) report the dated basalts to be slightly altered, which could have consequences for the accuracy and precision of their age.
The accuracy of the M-sequence age model of Ogg (2012) is ultimately dependent on the quality of available radioisotopic ages and cyclostratigraphic data close to or around stage boundaries from the Aptian to Oxfordian stages. New geochronological data from stage boundaries from the Late Jurassic to Early Cretaceous suggest that the age of the stage boundaries in this interval could be younger than used in the M-sequence model of Ogg (2012). For instance, Zhang et al. (2018) provided magnetostratigraphic data for the U–Pb ages of Midtkandal et al. (2016) in the Svalbard cores, which suggest that the age of the M0 (base of the Aptian) is 121–122 Ma rather than ∼126 Ma. Aguirre-Urreta et al. (2015) presented a high-precision U–Pb age of 127.24±0.03 Ma in the late Hauterivian (close to the base of the Barremian) in the Agrio Fm., Neuquén Basin, which Martinez et al. (2015) used to anchor cyclostratigraphic studies in the in Río Argos, Spain, and calculated an age for the base of the Hauterivian at 131.96±1 Ma and the base of the Barremian at 126.02±1 Ma. Aguirre-Urreta et al. (2017) later reported a U–Pb high-precision age at the early Hauterivian at 130.394±0.037 Ma, which is fairly close to that of Martinez et al. (2015) for the base of the Hauterivian. Therefore, new geochronological constraints in the Early Cretaceous render an apparent systematic offset ∼3–4 Myr younger than those used and predicted by the M-sequence age model of Ogg et al. (2012, 2016a). Incidentally, the data we present here for the J–K boundary and close to the KmTB display the same systematic offset (∼3–4 Myr) compared to the M-sequence model age of Ogg (2012) and Ogg et al. (2012).
In summary, the M-sequence age model for the Late Jurassic to Early Cretaceous stage boundaries is a creative solution to present numerical ages for stage boundaries with a clear lack of reliable radioisotopic ages. Nevertheless, recent geochronological developments in the Early Cretaceous show that some of the ages used to anchor the model are likely younger than previously accepted. Consequently, future updated versions of the M-sequence model are bound to incorporate these newer age constraints, and the critical overlap between the M-sequence model of Ogg (2012) and Mahoney et al. (2005) for the age of the J–K boundary is likely to change. Be that as it may, reliable radioisotopic ages for the J–K boundary with high-resolution biostratigraphical markers and magnetostratigraphy in a single section are still lacking, but growing evidence points to a younger age of the J–K boundary as well as other stage boundaries in the Late Jurassic and Early Cretaceous.
The age of the J–K boundary has been controversial and difficult to determine for the past decades. Our data presented here seem to restrict the J–K boundary from 140.7–140.9 Ma. This interval nevertheless carries uncertainty due to statistical interpolation of the age–depth modeling and estimated sedimentation rate. Our geochronology highlights the problem of using the FO and LO of key taxa between the studied sections. We suggest that this might be because of (1) different degrees of sample density, (2) insufficient frequency of taxa, (3) preservation of the geological record, and (4) environmental–depositional differences. Nevertheless, our data impose certain constraints for a J–K boundary age at ∼145 Ma. For instance, the late Tithonian assemblage of Crassicollaria parvula and Crassicollaria colomi and the FO of Umbria granulosa granulosa have an age of 141.31±0.56 Ma. The FO of R. asper is at 140.60±0.4 Ma, and the Virgatosphinctes andesensis zone FO is at 147.112±0.078 Ma, which calls for a revision of the age of the J–K boundary.
We are unable to precisely define the age of the J–K boundary, mainly because the biostratigraphy does not allow for the same temporal resolution as the geochronological methods used. Nevertheless, it is essential to recognize that the Las Loicas and Mazatepec sections are unique since they contain datable horizons close to or around the J–K boundary. Therefore, our U–Pb dates from these two sections, despite the discussed limitations, provide evidence for a younger numerical age of the J–K boundary.
All data are provided in the Supplement.
The supplement related to this article is available online at: https://doi.org/10.5194/se-10-1-2019-supplement.
The authors declare that they have no conflict of interest.
This paper is dedicated to the memory of Márcio Pimentel, who
unfortunately passed away during the reviewing process of this paper.
Márcio Pimentel was a champion of isotope geochemistry and geochronology in
Brazil and played a vital role in the development of personnel and
analytical capabilities of the field in Brazil during the 1990s and early
2000s. His passing is a great loss to the community, and his presence will
be sorely missed. The authors would like to thank reviewers William Wimbledon,
József Pálfy, and Bruno Galbrun for their valuable input, especially the editor Silvia Gardin for
the thorough and careful comments that enormously improved this
study. Luis Lena would like to thank CAPES (under project 1130-13-7) and the
University of Geneva for financial support. Marcio Pimentel thanks CNPq for
financial aid under project 470608/2013-7. Sam Bowring (MIT) is kindly
acknowledged for his support during the initial stages of the project. Adam Curry and Gregor Weber are warmly thanked for their help with the sample
from Mexico. This is contribution R-262 of the Instituto de Estudios Andinos
Don Pablo Groeber. Rafael López-Martínez was funded under grant PAPIIT IA103518.
Edited by: Silvia Gardin
Reviewed by: József Pálfy, William Wimbledon,
and Bruno Galbrun
Aguirre-Urreta, B., Rawson, P. F., Concheyro, G. A., Bown, P. R., and Ottone, E. G.: Lower Cretaceous (Berriasian-Aptian) biostratigraphy of the Neuquén Basin, Argentina, Geol. Soc. Sp., 252, 57–81, doi.org/10.1144/GSL.SP.2005.252.01.04, 2005.
Aguirre-Urreta, B., Vennari, V. V., Lescano, M., Naipauer, M., Concheyro, A., and Ramos, V. A.: Bioestratigrafía y geocronología de alta resolución de la Formación Vaca Muerta, cuenca Neuquina. IX Congreso de Exploración y Desarrollo de Hidrocarburos (Mendoza), Trabajos Técnicos II, 245–268, 2014.
Aguirre-Urreta, B., Lescano, M., Schmitz, M. D., Tunik, M., Concheyro, A., Rawson, P. F., and Ramos, V. A.: Filling the gap: new precise Early Cretaceous radioisotopic ages from the Andes, Geol. Mag., 152, 557–564, https://doi.org/10.1017/S001675681400082X, 2015.
Aguirre-Urreta, B., Schmitz, M., Lescano, M., Tunik, M., Rawson, P. F., Concheyro, A., Buhler, M., and Ramos, V. A.: A high precision U–Pb radioisotopic age for the Agrio Formation, Neuquén Basin, Argentina: Implications for the chronology of the Hauterivian Stage, Cretac. Res., 75, 193–204, https://doi.org/10.1016/j.cretres.2017.03.027, 2017.
Bakhmutov, V. G., Halásová, E., Ivanova, D. K., Józsa, Š., Reháková, D., and Wimbledon, W. A. P.: Biostratigraphy and magnetostratigraphy of the uppermost Tithonian–lower Berriasian in the Theodosia area of Crimea (Southern Ukraine), Geol. Q., 62, 197–236, https://doi.org/10.7306/gq.1404, 2018.
Baresel, B., Bucher, H., Brosse, M., Cordey, F., Guodun, K., and Schaltegger, U.: Precise age for the Permian-Triassic boundary in South China from high-precision U-Pb geochronology and Bayesian age-depth modeling, Solid Earth, 8, 361–378, https://doi.org/10.5194/se-8-361-2017, 2017.
Bown, P. R.: Early to Mid-Cretaceous calcareous nannoplankton from the northwest Pacific Ocean, Leg 198, Shatsky Rise, Proc. Ocean Drill. Program, Sci. Results, 198, 1–82, 2005.
Bralower, T. J., Monechi, S., and Thierstein, H. R.: Calcareous nannofossil zonation of the Jurassic-Cretaceous boundary interval and correlation with the geomagnetic polarity timescale, Mar. Micropaleontol., 14, 153–235, https://doi.org/10.1016/0377-8398(89)90035-2, 1989.
Bralower, T. J., Ludwig, K. R., and Obradovich, J. D.: Berriasian (Early Cretaceous) radiometric ages from the Grindstone Creek Section, Sacramento Valley, California, Earth Planet. Sc. Lett., 98, 62–73, https://doi.org/10.1016/0012-821X(90)90088-F, 1990.
Campa, M. F. and Coney, P. J.: Tectono-stratigraphic terranes and mineral resource distributions in Mexico, Can. J. Earth Sci., 20, 1040–1051, https://doi.org/10.1139/e83-094, 1983.
Casellato, C. E.: Calcareous nannofossil biostratigraphy of upper Callovian-lower Berriasian successions from the southern Alps, north Italy, Riv. Ital. di Paleontol. e Stratigr., 116, 357–404, 2010.
Channell, J. E. T., Erba, E., Nakanishi, M., and Tamaki, K.: Late Jurassic-early Cretaceous time scales and oceanic magnetic anomaly block models, in: Geochronology Time Scales and Global Stratigraphic Correlation, 54, 51–63, 1995.
Edwards, A.: A preparation technique for calcareous nannoplankton, Micropaleontology, 9, 103–104, 1963.
Elbra, T., Schnabl, P., Ková, K., Pruner, P., Kdýr, Š., Grabowski, J., Reháková, D., Svobodová, A., Frau, C., and Wimbledon, W. A. P.: Palaeo- and rock-magnetic investigations across Jurassic-Cretaceous boundary at St Bertrand's Spring, Drôme, France: applications to magnetostratigraphy, Stud. Geophys. Geod., 62, 323–338, https://doi.org/10.1007/s11200-016-8119-5, 2018.
Grabowski, J.: Magnetostratigraphy of the Jurassic/Cretaceous boundary interval in the Western Tethys and its correlations with other regions?: a review, Volumina Jurassica, 105–128, 2011.
Gradstein, F. M.: Introduction: In The Geologic Time Scale, 1–29, Elsevier, https://doi.org/10.1016/B978-0-444-59425-9.00001-9, 2012.
Gradstein, F. M., Agterberg, F. P., Ogg, J. G., Hardenbol, J., Veen, P. V, Thierry, J., and Huang, Z.: Comparison of Cretaceous Time Scales, Geochronol. Time Scales Glob. Stratigr. Correl., 54, 95–126, 1995.
Haslett, J. and Parnell, A.: A simple monotone process with application to radiocarbon-dated depth chronologies, J. R. Stat. Soc. C-Appl., 57, 399–418, https://doi.org/10.1111/j.1467-9876.2008.00623.x, 2008.
Hoedemaeker, P. J., Janssen, N. M. M., Casellato, C. E., Gardin, S., Reháková, D., and Jamrichová, M.: Jurassic/Cretaceous boundary in the Río Argos succession (Caravaca, SE Spain), Rev. Paleobiol., 35, 111–247, https://doi.org/10.5281/zenodo.51872, 2016.
Huang, C., Hinnov, L., Fischer, A. G., Grippo, A., and Herbert, T.: Astronomical tuning of the Aptian Stage from Italian reference sections, Geology, 38, 899–902, https://doi.org/10.1130/G31177.1, 2010a.
Huang, C., Hesselbo, S. P., and Hinnov, L.: Astrochronology of the late Jurassic Kimmeridge Clay (Dorset, England) and implications for Earth system processes, Earth Planet. Sc. Lett., 289, 242–255, doi.org/10.1016/j.epsl.2009.11.013, 2010b.
Kent, D. V. and Gradstein, F. M.: A Cretaceous and Jurassic geochronology, Geol. Soc. Am. Bull., 96, 1419–1427, 1985.
Kietzmann, D. A., Ambrosio, A. L., Suriano, J., Alonso, S., Gonz, F., Depine, G., and Repol, D.: The Vaca Muerta – Quintuco system (Tithonian – Valanginian) in the Neuquén Basin, Argentina: a view from the outcrops in the Chos Malal fold and thrust belt, Am. Assoc. Petr. Geol. B., 5, 743–771, https://doi.org/10.1306/02101615121, 2016.
Larson, R. L. and Hilde, T. W. C.: A revised time scale of magnetic reversals for the Early Cretaceous and Late Jurassic, J. Geophys. Res., 80, 2586, https://doi.org/10.1029/JB080i017p02586, 1975.
Legarreta, L. and Uliana, M. A. Jurassic-Cretaceous marine oscillations and geometry of back-arc basin fill, central Argentine Andes. International Association of Sedimentology, Special Publication, 12, 429–450, London, 1991.
Legarreta L. and Uliana, M. A.: The Jurassic succession in west-central Argentina: stratal patterns, sequences and paleogeographic evolution. Palaeogeogr. Palaeocl., 120, 303–330, https://doi.org/10.1016/0031-0182(95)00042-9, 1996.
López-Martínez, R., Barragán, R., Reháková, D., and Cobiella-Reguera, J. L.: Calpionellid distribution and microfacies across the Jurassic/Cretaceous boundary in western Cuba (Sierra de los Órganos), Geol. Carpathica, 64, 195–208, https://doi.org/10.2478/geoca-2013-0014, 2013a.
López-Martínez, R., Barragán, R., and Reháková, D.: The Jurassic/Cretaceous boundary in the Apulco area by means of calpionellids and calcareous dinoflagellates: An alternative to the classical Mazatepec section in eastern Mexico, J. South Am. Earth Sci., 47, 142–151, https://doi.org/10.1016/j.jsames.2013.07.009, 2013b.
López-Martínez, R., Barragán, R., Reháková, D., Martini, M., and de Antuñano, S. E.: Calpionellid biostratigraphy, U-Pb geochronology and microfacies of the Upper Jurassic-Lower Cretaceous Pimienta Formation (Tamazunchale, San Luis Potosí, central-eastern Mexico), Bol. la Soc. Geol. Mex., 67, 75–86, 2015.
López-Martínez, R., Aguirre-Urreta, B., Lescano, M., Concheyro, A., Vennari, V., and Ramos, V. A.: Tethyan calpionellids in the Neuquén Basin (Argentine Andes), their significance in defining the Jurassic/Cretaceous boundary and pathways for Tethyan-Eastern Pacific connections, J. South Am. Earth Sci., 78, 1–10, https://doi.org/10.1016/j.jsames.2017.06.007, 2017.
Lowrie, W. and Ogg, J. G.: A magnetic polarity time scale for the Early Cretaceous and Late Jurassic, Earth Planet. Sc. Lett., 76, 341–349, 1985.
Lukeneder, A., Halásová, E., Kroh, A., Mayrhofer, S., Pruner, P., Reháková, D., Schnabl, P., Sprovieri, M., and Wagreich, M.: High resolution stratigraphy of the Jurassic-Cretaceous boundary interval in the Gresten Klippenbelt (Austria), Geol. Carpathica, 61, 365–381, https://doi.org/10.2478/v10096-010-0022-3, 2010.
Mahoney, J. J., Duncan, R. A., Tejada, M. L. G., Sager, W. W., and Bralower, T. J.: Jurassic-Cretaceous boundary age and mid-ocean-ridge-type mantle source for Shatsky Rise, Geology, 33, 185–188, https://doi.org/10.1130/G21378.1, 2005.
Malinverno, A., Hildebrandt, J., Tominaga, M., and Channell, J. E. T.: M-sequence geomagnetic polarity time scale (MHTC12) that steadies global spreading rates and incorporates astrochronology constraints, J. Geophys. Res.-Sol. Ea., 117, 1–17, https://doi.org/10.1029/2012JB009260, 2012.
Martinez, M., Deconinck, J. F., Pellenard, P., Riquier, L., Company, M., Reboulet, S., and Moiroud, M.: Astrochronology of the Valanginian-Hauterivian stages (Early Cretaceous): Chronological relationships between the Paraná-Etendeka large igneous province and the Weissert and the Faraoni events, Global Planet. Change, 131, 158–173, https://doi.org/10.1016/j.gloplacha.2015.06.001, 2015.
Midtkandal, I., Svensen, H. H., Planke, S., Corfu, F., Polteau, S., Torsvik, T. H., Faleide, J. I., Grundvåg, S. A., Selnes, H., Kürschner, W., and Olaussen, S.: The Aptian (Early Cretaceous) oceanic anoxic event (OAE1a) in Svalbard, Barents Sea, and the absolute age of the Barremian-Aptian boundary, Palaeogeogr. Palaeocl., 463, 126–135, https://doi.org/10.1016/j.palaeo.2016.09.023, 2016.
Muttoni, G., Visconti, A., Channell, J. E. T., Casellato, C. E., Maron, M., and Jadoul, F.: An expanded Tethyan Kimmeridgian magneto-biostratigraphy from the S'Adde section (Sardinia): Implications for the Jurassic timescale, Palaeogeogr. Palaeocl., 503, 90–101, https://doi.org/10.1016/j.palaeo.2018.04.019, 2018.
Ogg, J. G.: Geomagnetic Polarity Time Scale, in The Geologic Time Scale, 85–113, Elsevier, https://doi.org/10.1016/B978-0-444-59425-9.00005-6, 2012.
Ogg, J. G. and Lowrie, W.: Magnetostratigraphy of the Jurassic / Cretaceous boundary, Geology, 14, 547–550, 1986.
Ogg, J. G., Hasenyager, R. W., Wimbledon, W. A., Channell, J. E. T., and Bralower, T. J.: Magnetostratigraphy of the Jurassic-Cretaceous boundary interval-Tethyan and English faunal realms, Cretac. Res., 12, 455–482, https://doi.org/10.1016/0195-6671(91)90002-T, 1991.
Ogg, J. G., Hinnov, L. A., and Huang, C.: Cretaceous, In The Geological Time Scale., 793–853, Elsevier., https://doi.org/10.1016/B978-0-444-59425-9.00027-5, 2012.
Ogg, J. G., Ogg, G. M., and Gradstein, F. M.: Cretaceous, in A Concise Geologic Time Scale, 167–186, Elsevier, 2016a.
Ogg, J. G., Ogg, G. M., and Gradstein, F. M.: Jurassic, in A Concise Geologic Time Scale, 151–166, Elsevier, 2016b.
Ovtcharova, M., Goudemand, N., Hammer, Ø., Guodun, K., Cordey, F., Galfetti, T., Schaltegger, U., and Bucher, H.: Developing a strategy for accurate definition of a geological boundary through radio-isotopic and biochronological dating: The Early-Middle Triassic boundary (South China), Earth-Sci. Rev., 146, 65–76, https://doi.org/10.1016/j.earscirev.2015.03.006, 2015.
Padilla and Sánchez, R. J.: Evolución geológica del sureste mexicano desde el Mesozoico al presente en el contexto regional del Golfo de México, Boletín la Soc. Geológica Mex., 59, 19–42, https://doi.org/10.18268/BSGM2007v59n1a3, 2007.
Pálfy, J.: The quest for refined calibration of the Jurassic time-scale, Proc. Geol. Assoc., 119, 85–95, https://doi.org/10.1016/S0016-7878(08)80260-X, 2008.
Pálfy, J., Smith, P. L., and Mortensen, J. K.: A U – Pb and 40 Ar/39 Ar time scale for the Jurassic, Can. J. Earth Sci., 944, 923–944, 2000.
Parnell, A. C., Haslett, J., Allen, J. R. M., Buck, C. E., and Huntley, B.: A flexible approach to assessing synchroneity of past events using Bayesian reconstructions of sedimentation history, Quaternary Sci. Rev., 27, 1872–1885, https://doi.org/10.1016/j.quascirev.2008.07.009, 2008.
R Core Team: RStudio Team, RStudio: Integrated Development for R. RStudio, Inc., Boston, MA, available at: http://www.rstudio.com/ (last access: 21 June 2018), 2015.
Renne, P. R., Mundil, R., Balco, G., Min, K., and Ludwig, K. R.: Joint determination of 40K decay constants and 40Ar*/40K for the Fish Canyon sanidine standard, and improved accuracy for 40Ar/39Ar geochronology, Geochim. Cosmochim. Ac., 74, 5349–5367, https://doi.org/10.1016/j.gca.2010.06.017, 2010.
Riccardi, A. C.: The marine Jurassic of Argentina?: a biostratigraphic framework, Episodes – Newsmag. Int. Union Geol. Sci., September, 326–335, 2008.
Riccardi, A. C.: Remarks on the Tithonian-Berriasian ammonite biostratigraphy of west central Argentina, Vol. Jurassica, XIII, 23–52, https://doi.org/10.5604/17313708, 2015.
Sager, W. W.: What built Shatsky Rise, a mantle plume or ridge tectonics?, Geol. Soc. Am. Spec. Pap., 388, 721–733, https://doi.org/10.1130/2005.2388(41), 2005.
Salvador, A.: Late Triassic- Jurassic paleogeography and origin of Gulf of Mexico basin, Am. Assoc. Petr. Geol. B., 63, 419–451, https://doi.org/10.1306/2F917F19-16CE-11D7-8645000102C1865D, 1987.
Selby, D., Mutterlose, J., and Condon, D. J.: U-Pb and Re-Os geochronology of the Aptian/Albian and Cenomanian/Turonian stage boundaries: Implications for timescale calibration, osmium isotope seawater composition and Re-Os systematics in organic-rich sediments, Chem. Geol., 265, 394–409, https://doi.org/10.1016/j.chemgeo.2009.05.005, 2009.
Suter, M.: Tectonics of the external part of the Sierra Madre Oriental foreland thurst-and-fold belt between xilitla and the Moctezuma river (Hidalso and San Luis Potose States), Rev. Mex. ciencias geológicas, 4, 19–31, 1980.
Svobodová, A. and Košťák, M.: Calcareous nannofossils of the Jurassic/Cretaceous boundary strata in the Puerto Escaño section (southern Spain)-biostratigraphy and palaeoecology, Czech Acad. Sci. v.v.i., Rozvojová, 67, 0–6, https://doi.org/10.1515/geoca-2016-0015, 2016.
Tamaki, K. and Larson, R. L.: The Mesozoic tectonic history of the Magellan microplate in the western central Pacific, J. Geophys. Res., 93, 2857–2874, https://doi.org/10.1029/JB093iB04p02857, 1988.
Tripathy, G. R., Hannah, J. L., and Stein, H. J.: Refining the Jurassic-Cretaceous boundary: Re-Os geochronology and depositional environment of Upper Jurassic shales from the Norwegian Sea, Palaeogeogr. Palaeocl., 503, 13–25, https://doi.org/10.1016/j.palaeo.2018.05.005, 2018.
Vennari, V. V., Lescano, M., Naipauer, M., Aguirre-urreta, B., Concheyro, A., Schaltegger, U., Armstrong, R., Pimentel, M., and Ramos, V. A.: New constraints on the Jurassic-Cretaceous boundary in the High Andes using high-precision U-Pb data, Gondwana Res., 26, 374–385, https://doi.org/10.1016/j.gr.2013.07.005, 2014.
Vennari, V. V.: Tithonian ammonoids (Cephalopoda, Ammonoidea) from the Vaca Muerta Formation, Neuquén Basin, West-Central Argentina, Palaeontogr. Abt. A, 306, 85–165, https://doi.org/10.1127/pala/306/2016/85, 2016.
Wimbledon, W. A. P.: Developments with fixing a Tithonian/Berriasian (J/K) boundary, Vol. Jurassica, 15, 181–186, https://doi.org/10.5604/01.3001.0010.7467, 2017.
Wimbledon, W. A. P., Casellato, C. E., Reháková, D., Bulot, L. G., Erba, E., Gardin, S., Verreussel, R. M. C. H., Munsterman, D. K., and Hunt, C. O.: Fixing a basal Berriasian and Jurassic/Cretaceous (J/K) boundary – Is there perhaps some light at the end of the tunnel?, Riv. Ital. di Paleontol. e Stratigr., 117, 295–307, 2011.
Wimbledon, W. A. P., Reháková, D., Pszczółkowski, A., Casellato, C. E., Halásová, E., Frau, C., Bulot, L. G., Grabowski, J., Sobień, K., Pruner, P., Schnabl, P., and Čížková, K.: An account of the bio- and magnetostratigraphy of the Upper Tithonian–Lower Berriasian interval at Le Chouet, Drôme (SE France), Geol. Carpathica, 64, 437–460, https://doi.org/10.2478/geoca-2013-0030, 2013.
Wotzlaw, J.-F., Brack, P., and Storck, J.-C.: High-resolution stratigraphy and zircon U-Pb geochronology of the Middle Triassic Buchenstein Formation (Dolomites, northern Italy): precession-forcing of hemipelagic carbonate sedimentation and calibration of the Anisian-Ladinian boundary interval, J. Geol. Soc. London., https://doi.org/10.1144/jgs2017-052, 2017.
Zhang, Y. W., Minguez, Ogg, D., and Olaussen, J. G.: Magnetostratigraphy across u-pb-dated horizons in svalbard boreholes to partly resolve the debated age of barremian/aptian boundary, Geol. Soc. Am., 50, https://doi.org/10.1130/abs/2018AM-323068, 2018.